
Зимовка и терморегуляция пчелосемьи
Андрес Раава
Раздел 1.9 книги А. Раава "Мой улей", дополненный автором.
Осенью, когда температура вне улья становится ниже +10°С, пчелы начинают объединяться в зимний клуб. Клуб имеет форму эллипсоида или растянутого по вертикали шара.. В ядре клуба пчелы сидят неплотно и могут свободно двигаться. Эти пчелы, изменяя интенсивность своего движения, поддерживают температуру ядра в пределах +14...+30°С. Ядро покрывается оболочкой, состоящей из плотно сидящих пчел. В зависимости от температуры гнездового пространства, пчелы регулируют толщину слоя облочки от 2,5 до 7 см. Так пчелосемья старается поддерживать стабильную температуру (+8°С) на поверхности клуба. У пчел, объединенных в клуб, замедляется ритм жизни и протекание физиологических процессов. На основной обмен веществ (метаболизм) зимний клуб пчел расходует, в среднем, приблизительно 1 грамм меда в час, продуцируя в тоже время приблизительно 2,8 Ккал тепла, 1,5 грамма углекислого газа и 0,5 грамма водяного пара. В зависимости от изменений внеульевой температуры, которые неизбежно вызывают изменения температуры в гнездовом пространстве, зимующий клуб пчел изменяет толщину своей оболочки и увеличивает потребление корма (меда). Приблизительно так описывается зимовка пчелосемьи в книгах, которые знакомят читателей с пчелосемьей, пчелами, их поведением и способностью регулировать температуру.
Что и как происходить в улье зимой, помог мне разгадать снимок, который я сделал с погибшей пчелосемьи.
В начале лета 1986 года я раскрыл новые основы содержания пчелосемей и, с учетом раннего опыта, мне стало ясно, что пчелосемья способна подготовить себя к зимовке самостоятельно гораздо лучше, чем при помощи и вмешательства человека. Семье надо только дать возможность для этого. Правильность моей теории подтверждает факт, что в нынешнем виде пчелосемья существует в экосистеме Земли уже 65 миллионов лет и за все это время прекрасно справлялась с зимовкой. Человек начал готовить пчелосемью к зимовке и "обматывать одеялом" только в течение пары последних столетий. Также в пчеловодной литературе утверждается, что пчелосемья холода не боится и приводится множество примеров, подтверждающих это.
Осенью 1986 года я начал готовить пчелосемьи к зимовке по одному принципу - обеспечивал их только кормом. Но из-за проведенных летом экспериментов, одна пчелосемья осталась зимовать в стадии замены ульевых рамок. Причиной такого состояния пчелосемьи была ее низкая скорость роста из-за малого веса. Необходимым кормовым запасом для зимовки семья была обеспечена.
Зима 1986/87 года была очень суровой, внешняя среднесуточная температура в декабре в течение нескольких недель колебалась в пределах минус 15...30°С и в начале января 1987 года эта семья погибла. Мне было ясно, что семья погибла не из-за недостатка корма в улье. По моему мнению, причиной гибели семьи могла быть только замена местонахождения ульевых рамок во время обеспечивания ее кормовыми запасами и попытка вывести из улья стандартные ульевые рамки, вследствие чего, были вызваны нарушения формирования зимнего клуба.
Для того, чтобы убедиться в своих предположениях, я решил при первой возможности осмотреть погибшую семью. Когда между крепкими морозами стало немного теплее (-7...-9°С) я это сделал и запечатлел осмотр, шаг за шагом, на фотопленку. В ходе осмотра оказалось, что я правильно угадал причину гибели семьи. Но я увидел ещё много интересного, чего нельзя увидеть после растаивания улья с погибшей пчелосемьей.
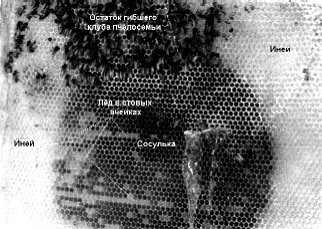 При осмотре фотографии погибшей семьи (см. фото), меня удивил не столько находящийся в улье и на сотах иней, сколько сосулька, находящаяся немного ниже остатка клуба. Сосулька может образоваться только тогда, когда в гнездовом пространстве есть одновременно тепло и мороз. Это означает, что сосулька могла образоваться только во время жизни пчелосемьи. Но в это же время в улье должен был быть и мороз, чтобы вода замерзла.
При осмотре фотографии погибшей семьи (см. фото), меня удивил не столько находящийся в улье и на сотах иней, сколько сосулька, находящаяся немного ниже остатка клуба. Сосулька может образоваться только тогда, когда в гнездовом пространстве есть одновременно тепло и мороз. Это означает, что сосулька могла образоваться только во время жизни пчелосемьи. Но в это же время в улье должен был быть и мороз, чтобы вода замерзла.
Меня стал мучить вопрос: что же, все-таки, защищает пчелосемью от холода и, что происходит в улье зимой во время зимовки пчелосемьи?
Этот вопрос мучил меня несколько недель, пока мне внезапно в голову не пришла мысль - а что, если покрыть поверхность клуба пчелосемьи слоем неподвижного воздуха толщиной около 10 мм (максимальный размер "пространства Лангстрота")? Я сразу сделал необходимые теплотехнические расчеты. За основу для расчетов взял вышеприведенные данные по основному обмену веществ клуба пчелосемьи и диаметру шарообразного зимнего клуба пчелосемьи - 20 см. Верность теплотехнических расчетов я проверил несколькими методами, в том числе, методом на основе закона Ома по эквивалентной схеме двухполярного источника напряжения и с научными сотрудниками теплотехнической кафедры Таллиннского Политехнического Института.
На основе расчетов и раннего опыта я сразу создал новую теорию о зимовке пчелосемьи, которая доказывает, что:
- зимующий клуб пчелосемьи можно защитить окружающим его неподвижным воздухом от любых отрицательных значений температуры внешней среды тем количеством тепла, которое выделяется клубом в ходе основного обмена веществ (метаболизма);
- находящийся в неподвижным воздухе клуб пчелосемьи защищает от влаги - мороз, и от мороза, в свою очередь, выделяемая клубом, влага;
- клуб пчелосемьи является теплокровным существом, способным регулировать интенсивность обмена веществ для продуцирования тепла, необходимого для поддержания биологически предназначенной температуры +8°С на поверхности своего тела (клуба) и для компенсирования тепловых потерь с поверхности клуба;
- понятие "внешняя среда" для клуба (пчелосемьи) начинается с поверхности клуба (тела) пчелосемьи, а не с внешней поверхности улья.
Что же в действительности происходит в улье при зимовке пчелосемьи?
Когда температура внутренней поверхности гнезда падает ниже, чем температура поверхности клуба пчелосемьи (+8°С), которую клуб поддерживает стабильной, начинается циркуляция воздуха в гнездовом пространстве. Циркуляция воздуха вызывается разницей температур на поверхности клуба пчелосемьи и на внутренней стене гнезда. Интенсивность циркуляций воздуха зависит от расстояния между поверхностью клуба пчелосемьи и внутренной стеной гнезда и от величины разницы температур на поверхности клуба и на поверхности внутренней стены гнезда. На циркуляцию воздуха не влияет то обстоятельство, что гнездовое пространство ниже уровня размещения клуба пчелосемьи закрыто или немного открыто. Также, выделяемое клубом тепло (около 2,8 Ккал в час) в ходе основного обмена веществ, не может препятствовать снижению температуры на внутренней стене гнезда ниже +8°С и проникновению в гнездовое пространство мороза. Чтобы понять как мало количество тепла, выделяемое клубом, надо изготовить, примерно из 3 грамм пчелиного воска, свечу и дать ей гореть в течение одного часа. Тогда из неё выделяется примерно столько же тепла, сколько клуб выделяет в течение часа вследствие основного обмена веществ. Становится ясно, что невозможно различным утеплением стен улья препятствовать снижению температуры внутренней поверхности гнезда ниже температуры поверхности клуба пчелосемьи и проникновению отрицательной температуры в гнездовое пространство. Причина очень простая - пчелосемья начинает продуцировать дополнительно тепло (кроме продуцированного в ходе основного обмена веществ) только тогда, когда проникнувшая в гнездовое пространство температура отрицательна по отношению к температуре поверхности клуба, и, которая вызывает такие потери тепла с поверхности клуба, что температура на его поверхности падает ниже +8°С. Чтобы такая ситуация возникла раньше, чем температура на внутренней стене гнезда упадет ниже 0°С, расстояние от поверхности клуба пчелосемьи до внутренней стены гнезда должно быть очень большим. Это расстояние, в свою очередь, зависит от длины улочек (гнездовой рамки) и в некоторой мере от ширины гнезда. Чем меньше длина применяемых ульевых рамок превышает диаметр клуба пчелосемьи, тем меньше циркуляция воздуха в гнездовым пространстве и, вместе с тем, теплопередача посредством воздуха с поверхности клуба на внутреннюю стену гнезда. Если длина ульевой рамки такая, что расстояние от поверхности клуба пчелосемьи до внутренней стенки гнезда остается меньше 5 см, тогда циркуляция воздуха вообще не возникнет. Это происходит из-за физических свойств воздуха. В вертикальном слое воздуха толщиной меньше 5 см не может возникнуть двухстороннее движение воздуха (с поверхности клуба вверх и со стенки улья вниз) за счет силы трения между молекулами воздуха. Эта сила трения не допускает двухстороннего движения в слое воздуха толщиной 5 см и меньше, при любых разницах температур на поверхности такого слоя воздуха, воздух остается неподвижным и тепловые потери с поверхности клуба (теплообмен) будут только излучательными. Когда температура на внутренней стене гнезда падает ниже 0°С, между поверхностью клуба и стеной гнезда образуется скалярное поле температуры 0°С (скаляр - значение, определяемое только одним числовым значением). Разрез скалярного поля называют изотермом. После образования скалярного поля 0°С, начинается интенсивное конденсирование водяных паров, находящихся в гнездовом пространстве, и на частях гнезда и на сотах при отрицательных температурах образуется иней. Вследствие этого процесса воздух в гнездовом пространстве вокруг клуба высушивается и выпадающий иней начинает препятствовать возможной локальной циркуляции воздуха в отдельных частях гнездового пространства, где расстояние от поверхности клуба до стены гнезда превышает 5 см.
В сухом неподвижном воздухе, выделяемое клубом в ходе основного обмена веществ тепло, останавливает приближение скалярного поля 0°С на расстояние около 10 мм от поверхности клуба (максимальный размер "пространства Лангстроота"). В зависимости от расхода запасного вещества (меда) внутри клуба, клуб передвигается вверх, по естественному направлению тепла, приближаясь к скалярному полю 0°С и инею над клубом. Вследствие этого иней сверху клуба тает и, образуемая вода стекает по поверхности скалярного поля 0°С вниз. Протекающая вниз вода, попадая снова в зону отрицательных температур, образует на сотах под клубом сосульки. Находящийся постоянно в таком состоянии клуб пчелосемьи выдержит, при минимальной затрате энергии, любую внеульевую отрицательную температуру (до 0°К или -273,15°С) до тех пор, пока у клуба хватает запаса энергии и, у образующих клуб пчелосемьи особей, хватит ресурса обмена веществ, т.е. функционального резерва. Особи пчелосемьи, у которых кончается функциональный резерв, гибнут, выпадают из клуба и образуют на дне гнезда подмор. Температура поверхности клуба +8°С является биологически предназначенной и пчелы не могут сами ее изменять. Для поддержания биологически предназначенной температуры на поверхности клуба во время зимней спячки, клуб пчелосемьи продуцирует тепло периодично. Когда температура внутренней части клуба, за счет тепловых потерь, понижается до +14°С, центральная часть клуба начинает продуцировать тепло. Когда, за счет продуцируемого клубом тепла, температура центра клуба поднимается до +30°С, клуб прекращает продуцирование тепла и, регулированием теплопроводимости покровного слоя клуба, поддерживает температуру на поверхности клуба +8°С. Когда температура внутри клуба снова понижается до +14°С, цикл повторяется. Чем больше тепловые потери с поверхности клуба, тем чаще клубу приходится продуцировать тепло, расходовать мед и функциональный ресурс метаболизма особей пчелосемьи, образуюющих клуб. Если пчелосемья не способна по какой-либо причине сохранять биологически предназначенную температуру поверхности клуба, она начинает погибать. Первыми гибнут рабочие пчелы на поверхности клуба и выпадают из клуба. Если клубу не удается приостановить этот процесс, гибнет весь клуб. Температура тела ниже +8°С является летальной (смертельной) для любой особи пчелосемьи, независимо от того, находится она в клубе или вне клуба. Ни одна особь пчелосемьи вне клуба не способна регулировать теплопродуктивность своего тела. Этим свойством обладает только клуб пчелосемьи, поэтому нельзя говорить, что пчелы способны регулировать (изменять) температуру гнезда. Пчелосемья (клуб) способна только регулировать интенсивность обмена веществ для продуцирования тепла с целью сохранения биологически предназначенной температуры клуба, т.е. клуб имеет способность терморегуляции. Отделяющаяся от клуба пчелосемьи особь, является хладнокровной в пределах температуры своего тела от +8...70°С (по Г.Ф. Таранову).
Если температура на поверхности внутренней стены улья поднимается выше 0°С, с гнездового пространства пропадает скалярное поле 0°С, образовавшиеся иней и лед начинают медленно таять и эта вода сливается в пустые сотовые ячейки, где образует необходимый запас воды для весеннего развития пчелосемьи. Такой запас воды незначительно влияет на влажность воздуха гнездового пространства из-за маленькой поверхности испарения и сравнительно низкой температуры воды.
Когда температура окружающей клуба среды поднимается выше +8°С, система терморегуляции клуба переключает пчелосемью в режим активной жизнедеятельности и пчеломатка начинает класть яйца, т.е. восстанавливается физиологическая регенерация пчелосемьи. Но этого может и не произойти, если функциональный резерв особей пчелосемьи или запасы энергии (меда) являются недостаточными для обеспечения проведения необходимых физиологических процессов по восстановлению и сохранению жизнеспособности пчелосемьи.
Но подъем температуры на внутренней стене гнезда выше 0°С не всегда вызывается подъемом внеульевой температуры выше 0°С. Такой подъем температуры на внутренней стене улья может быть вызван самой пчелосемьей при значительных колебаниях отрицательной внеульевой температуры, например при резком подъеме температуры от -20°С до -1°С, в случае, если теплоемкость улья большая, наружные стены хорошо утепленные и расстояние от поверхности клуба до внутренней стены улья превышает 5 см. В таких условиях пчелосемье приходится, при внеульевой температуре -20°С, вырабатывать значительное количество тепла для отопления воздуха в гнездовом пространстве. При резком повышений внеульевой температуры, высокая теплоемкость и утепление стенок улья, препятствует обогреву стен улья и выходу продуцируемого клубом тепла из гнездового пространства улья и пчелосемья нагревает воздух в гнездовом пространстве выше +8°С и поэтому переходит в режим активной жизнедеятельности, независимо от внеульевых условий.
Биологически предназначенная температура внутри клуба пчелосемьи (температура расплода), во время периода активной жизнедеятельности, является +34...36°С и пчелосемья сама не может регулировать (изменять) эту температуры. Для поддержания биологически предназначенной температуры во время периода активной жизнедеятельности, пчелосемья постоянно продуцирует тепло внутри клуба, изменяя интенсивность метаболизма. Интенсивность метаболизма регулируется тепловыми потерями с поверхности покровного слоя расплодного клуба, который образуется из плотно сидящих рабочих пчел. Если тепловые потери не удается покрыть продуцированием тепла, пчелосемья увеличивает толщины покровного слоя и, если это тоже не помогает, клуб уменьшает свой объем и, тем самым, площадь поверхности тела. Вследствие этого тепловые потери уменьшаются и клуб может сохранять необходимую температуру внутри клуба. Расплод, который, вследствие уменьшения диаметра (поверхности) клуба, попадает в покровный слой или за его пределы, гибнет.
В остальном, все факторы, определяющие терморегуляцию пчелосемьи во время периода активной жизнедеятельности, являются аналогичными зимним. Только количество тепла и температура тепла являются иными. Чтобы избежать лишних тепловых потерь расплодного клуба и бессмысленных расходов энергии в виде корма или меда, для компенсации тепловых потерь или перегрева во время жары, целесообразно, чтобы расстояние от поверхности покровного слоя клуба до внутренней стены улья было меньше 5-ти см. При этом теплоемкость улья и тепловое сопротивление стен улья должны быть как можно меньшими. Это обеспечивает получения клубом пчелосемьи объективную информацию о внеульевых условиях и об их изменениях.
АНДРЕС РААВА
65001 Миссо, Выруский уезд, Эстония
Тел.: 372 52 96 193
E-mail: info@pchelopas.ru